Функции и рост подземных органов растений
Корневые
системы имеют решающее значение для поглощения растениями питательных веществ и
влаги. Их формирование, как и образование надземных частей, протекает под
комплексным влиянием среды и генетических факторов. Однако
следует отметить, что до настоящего времени влияние внешних и внутренних
факторов на надземные части растений изучено лучше, чем на подземные.
Особенно слабо исследованы корневые системы тех культур, подземные части
которых не представляют товарной ценности.
До недавнего
времени основное внимание уделяли изучению экологических реакций и физиологии
надземных частей растений, роль же подземных рассматривалась лишь с точки
зрения закрепления растений в почве, обеспечения их водой и питательными веществами
или накопления и хранения запасных веществ. Также установлена физиологическая и
экологическая роль подземных органов для целого растения.
Подземные
органы оказывают влияние на рост и развитие всего растения, особенно с точки
зрения сопротивляемости засухе, всоким и низким
температурам, которые воздействуют на комплексное развитие и созревание
генеративных органов. Большое значение имеет образование ауксинов,
гиббереллинов, цитокининов, витаминов и других
веществ, которые через взаимодействие надземной и подземной частей растения
влияют на метаболизм и рост, на ускорение или торможение этих процессов. Не
менее важное значение имеет передвижение ассимилятов
и витаминов от надземных частей к подземным или их
транспорт в обратном направлении.
Влияние факторов вегетации на корневую систему
На подземные
части растений, как и на надземные, существенное влияние оказывают
вегетационные факторы, действующие непосредственно или косвенно, через
надземную массу.
Прямое
действие света на корни в нормальных условиях не проявляется, однако установлено,
что их формирование ослабевает при снижении освещенности надземных, фотосинтетически активных частей
как однолетних полевых культур, так и многолетних травостоев.
Рост корней
начинается при определенной температуре окружающей среды и ускоряется по мере
ее повышения до определенного, оптимального для данной культуры предела, а
затем замедляется. При наступлении температурного максимума рост подземных
органов прекращается. Корни, как правило, растут при более низких температурах,
чем надземные части, причем получены данные о росте корневой системы
многих растений, например трав, даже при температуре ниже нуля.
* Очевидно
речь идет об отрицательных температурах воздуха, а не почвы, причем в весьма
ограниченный отрезок времени (Прим.ред).
К высоким
температурам корни чувствительнее, чем надземные части растений.
Вода –
решающий фактор в развитии подземных органов, однако при определенных условиях
влажность влияет на корни слабее, чем на надземные части. На основании многочисленных
опытов с зерновыми установлено, что при орошении посевов масса корней в
поверхностном слое почвы увеличивается, а их проникание в более глубокие горизонты
ослабевает. У неорошаемых культур наибольшая масса корней развивается перед или в период кущения, тогда, как при орошении
нарастание корневой массы происходит и в более поздние сроки. Такая же
закономерность проявляется и по отношению к атмосферным осадкам. В почвах
засушливых областей корни растений прорастают на большую глубину, чем во влажных. Это влияние имеет свой предел: они не проникают
в тот слой почвенного профиля, влажность которого ниже влажности завядания.
Рост и
ветвление корней подчиняются общей закономерности: при более высокой влажности
субстрата длина корней снижается, а их разветвление усиливается. У растений с
хорошо развитым главным корнем отмечена различная реакция всей корневой системы
на различные условия увлажнения: при высоком содержании воды развивается густая
сеть разветвленных корней, а при высыхании почвы идет рост главного корня в
длину. Только с дальнейшим снижением влажности развитие корневой системы
тормозится, поэтому при сильном высыхании почвы в летнее время рост корней многолетних
кормовых культур ослабевает. Следовательно, рост корней наиболее сильно
колеблется в поверхностном слое почвы, где часто изменяются влажность и
температура, тогда как в глубоких горизонтах почвенного профиля динамика роста
более равномерна.
Из смеси
газов, которую представляет собой воздух, на подземные органы влияют прежде всего углекислый газ и кислорорд.
При высокой концентрации СО2 в воздухе почвы
рост корней тормозится, однако токсические концентрации СО2 в
культурных почвах встречаются лишь в исключительных случаях.
Питательные
вещества, особенно отдельные элементы, существенно влияют на рост и общую массу
подземных частей. Азот как основной элемент питания в жизни растений в
наибольшей степени определяет рост подземных органов большинства культур, хотя
его влияние на надземные части более существенно.
Роль фосфора в
большинстве случаев значительно слабее, причем сказывается одинаково на
развитии и подземных и надземных органов. Что касается его влияния на морфологию
корня, то следует отметить, что фосфор сдерживает рост корней большинства
растений в длину и усиливает ветвление.
Действие калия
идентично действию фосфора. В плохо обеспеченной калием среде происходит
дегенерация корневой системы. У ярового ячменя, например, при недостатке калия
полное омертвление корневых тканей отмечается уже в фазе молочной спелости.
Кальций и
магний способствуют большему ветвлению корней, при их недостатке тормозится
деление клеток в верхушечной меристеме и в результате снижается рост корней в
длину.
В мировой
литературе данных о влиянии микроэлементов на корневую систему мало, причем
часто они противоречивы. Однако значение микроэлементов для подземных органов
не менее велико, чем для надземных: как абсолютный дефицит, так и избыточное их
количество играют в развитии корней отрицательную роль.
Влияние
генетических факторов на формирование корней
На формировании
подземных органов растений заметно отражаются как внешние, так и внутренние
(генетические) факторы. Рост отдельных видов культур, а нередко и отдельных
сортов, их обусловленная генетически отзывчивость на внешние факторы часто
бывают различными. Поэтому некоторые генетически закрепленные признаки
подземных органов (например, число зародышевых корешков у зерновых) часто
используется при выведении новых сортов. В селекции (выведении новых сортов)
большинства культур считается целесообразно использовать те сорта, у которых
мощная корневая система – это генетически обусловленный признак.
Рост корней в полевых условиях
В предыдущей
главе были отдельно рассмотрены внешние (вегетационные) и внутренние
(генетические) факторы, влияющие на рост корневой системы. Однако в полевых
условиях на корни воздействует целый комплекс различных факторов, как
естественных, так и созданных деятельностью человека. Предположение о том, что
выход товарной продукции зависит от корневой массы, относится не ко всем
культурам, и это всегда необходимо иметь в виду (рис. ). Высокоактивная, но относительно
небольшая корневая система, которую часто развивают растения на гидропонике, в
сочетании с оптимальными для деятельности подземных и надземных органов
условиями является основой развития высокопродуктивной культуры, значительного
урожая биомассы и товарной продукции. Другие закономерности отмечаются для
многолетних бобовых трав (табл. ), где налицо прямая зависимость формирования
надземной массы от массы корней.
Число корней,
их общие длина и поверхность очень велики. Особенно это относится к зерновым
культурам, которые имеют особенно много корневых волосков. В поверхностном слое
почвы глубиной до
Размещение
подземных частей растений в почве обусловливается в первую очередь комплексом
окружающей среды, т.е. условиями произрастания и свойствами почвы. Основная
часть корневой системы культурных растений расположена в наиболее плодородной
части почвенного профиля. Глубина проникания и объем корней в большинстве случаев
меняются в зависимости от глубины пахотного слоя, в котором расположено 70-90%
их массы.
Колебания
общего количества и массы корней в каждом конкретном случае происходят в
основном под влиянием погоды в течение вегетации, тогда как условия произрастания
и тип почвы играют меньшую роль.
Содержание в
почве гумуса положительно сказывается на развитии корневой системы. Даже
глубоко расположенные в почвенном профиле слои и затеки гумуса обильнее
пронизываются корнями, чем более бедная гумусом среда.
Реакция
большинства пахотных почв, как правило, не оказывает отрицательного влияния на
корни растений. Изменения, которые в первую очередь появляются на кончиках
корней и корневых волосках, происходят только при экстремальной кислотности или
щелочности.
Однако реакция
почвы может косвенно отражаться на развитии подземных органов, например в
результате токсичности ионов алюминия, высвобождающихся при повышенной кислотности
почвы. Некоторые виды культурных растений под влиянием колеблющегося рН на разной глубине почвы развивают корни в ее
поверхностных слоях, это же отмечается и при высоком уровне грунтовых вод. Рост
корней люцерны полностью приостанавливается при рН
4,8. У большинства культур в почвах с возрастающей кислотностью сдерживается
ветвление корней, снижается количество корневых волосков, иногда отмечается
утолщение корневых окончаний и ослабление клеточных стенок.
Такое же
влияние оказывает щелочная реакция среды, особенно при недостатке влаги.
На молодые
корешки прорастающих семян отрицательно влияют специфические ионы и повышенные
концентрации солей в окружающей среде, а также высокие нормы минеральных
удобрений. В этих случаях под влиянием осмотического давления почвенного
раствора могут произойти разрыв клеток корней, их отмирание или заболевание.
Развитие
корней зависит от плотности почвы. Встречая на своем пути плотные слои почвы,
растущие корни проникают в глубину лишь по щелям и трещинам, оставшимся после
корней предшественника или обусловленным структурой почвы. Неблагоприятное
влияние чередования плотных и рыхлых слоев почвы хорошо заметно у сахарной
свеклы, у которой после предпосевной обработки, проведенной при повышенной
влажности почвы, обычно начинается ветвление корнеплодов.
Влияние агротехники на развитие подземных органов
Агротехнические
мероприятия, направленные на повышение количества товарной продукции, в большей
или меньшей степени отражаются на развитии всех органов растений, в том числе и
подземных.
Обработка
почвы в первую очередь положительно влияет на подземные органы при
неблагоприятных условиях произрастания. На плодородных почвах даже после
многолетних вспашек на глубину 10-
На почвах с
уплотненным подпахотным слоем или с разной степенью оглеения
обработка почвы может способствовать развитию и росту корней.
Отрицательное
влияние чередования культур на корневые системы сказывается
лишь на культурах, особенно чувствительных к предшественнику, или при
монокультуре (лен, клевер луговой). Причина этого заключается в накоплении в
почве веществ, тормозящих рост корней. Повреждение первичных корешков озимой
пшеницы происходит при высеве семян непосредственно после запашки соломы или
высокой стерни.
Умеренные
нормы азота, фосфора и калия, а также органических удобрений, как правило,
содействуют росту подземных органов всех культур. При этом наибольшее влияние
на формирование корней оказывают минеральные удобрения, особенно азотные. При
возрастающих нормах азотных удобрений урожай надземной массы повышается больше,
чем масса корней. Для корней максимальная эффективность азота была отмечена при
низких дозах внесения, тогда как для надземных частей – при высоких. В деталях
этих общих закономерностей проявляется существенная разница влияния культур и
сортов.
При внесении
возрастающих доз азота под клевер луговой снижалась продуктивность не только
надземной, но и подземной массы. Из микроэлементов массу корней обычно
увеличивает молибден, для других это не выявляется на статистически существенном
уровне. Суммарное внесение микроэлементов почти всегда дает прирост массы корней.
В мировой
литературе часто сообщается о поверхностном расположении корней многолетних трав,
интенсивно удобренных азотом. При выпасе скота растения с расположенными у
поверхности почвы и слабыми корнями легко выдергиваются, при этом повреждается
дернина.
Регуляторы
роста, применяемые для сокращения длины стебля у зерновых, иногда действуют на
рост корней положительно, иногда отрицательно. В данном случае большое влияние
оказывают вид культуры, сорт, доза и срок применения ростовых веществ.
Уборка и
удаление надземных частей растений или повреждение их ассимиляционной
поверхности приводят к ослаблению поступления ассимилятов
в подземные органы. Типичным примером служат многолетние травы: чем чаще
проводят укосы, тем меньше формируется корней и их рост снижается.
Фотосинтез в агроэкосистемах
Экосистемы,
питательные вещества, вода и другие факторы вегетации ,
подземные и надземные части растений служат основой и одновременно составной
частью фотосинтеза – самого важного процесса, создающего возможность
существования жизни на Земле. При фотосинтезе под влиянием комплекса факторов энергия
падающих солнечных лучей превращается в формы, используемые человеком:
продукты питания, одежду, топливо и т.д. Особое значение имеет солнечная
радиация в области длин волн 380-710 нм. Она называется ФАР (фотосинтетически активная радиация), поскольку почти вся ее
энергия может быть использована для фотосинтеза.
Как процесс
биохимический и биофизический фотосинтез в агроэкосистемах
не отличается от фотосинтеза в естественных экосистемах. Существенная разница в
количественных параметрах возникает в связи с тем, что в посевах изменяется
годовая ритмика фотосинтеза и другие параметры, направленные на то, чтобы
увеличить долю необходимого продукта (урожая).
Фотосинтез как диффузия СО2
В общем
понимании фотосинтез представляет собой абсорбцию углекислого газа (абсобция – поглощение с проникновением внутрь; адсорбция
– поглощение на поверхности). Из воздуха, окружающего растение, молекула СО2
должна поступить в хлоропласты, где она связывается соответствующим акцептором.
Этот перенос СО2 происходит в основном путем диффузии – передвижения
от мест с более высокой концентрацией к местам с более низкой концентрацией. В
последние десятилетия этого рода процессы изучались активно, и данные о
переносе СО2 теперь относятся к простым характеристикам фотосинтетического
аппарата, которые можно рассчитать на основе измерений, используя математические
формулы.
Скорость
диффузии, т.е. количество диффундирующего газа , тем больше, чем выше градиент
концентрации (разница в концентрации диффундирующего газа) и чем больше
поперечное сечение разреза, через который молекулы газа диффундируют.
Интенсивность
фотосинтеза, выраженную скоростью поглощения СО2
единицей площади в единицу времени, определяют прямым измерением при
одновременном выявлении концентрации СО2 в воздухе. Таким методом устанавливают
не только общее сопротивление диффузии, но и его компоненты. Молекулы из
окружающего лист воздуха проникают вначале через тонкий (десятки миллиметров)
слой почти неподвижного воздуха, соприкасающегося непосредственно с
поверхностью листа, преодолевая при этом сопротивление этого слоя. Далее через
устьица и межклетники и, наконец, через стенки клеток, плазмалемму, цитоплазму
и мембраны хлоропласта они попадают в зону, где осуществляется карбоксилирование. Оказываемое молекулам сопротивление
называют мезофильным, оно включает как собственно
транспортное (физическое) сопротивление, так и химическое, характеризующее
эффективность реакции карбоксилирования.
Рассмотрение
общего сопротивления переносу СО2 при
фотосинтезе в отдельных составных частях дает ценные сведения о месте действия
соответствующего фактора. При оценке влияния орошения, удобрений, регуляторов
роста и других факторов аграрного производства, такой анализ позволяет изучить
механизм воздействия отдельных частей и обобщить конкретные результаты.
Изменение устьичного сопротивления оказывает воздействие на устьица,
тогда как по колебаниям мезофильного сопротивления
можно судить об изменениях анатомической структуры или биохимических реакций.
Влияние внешних и внутренних факторов на фотосинтез
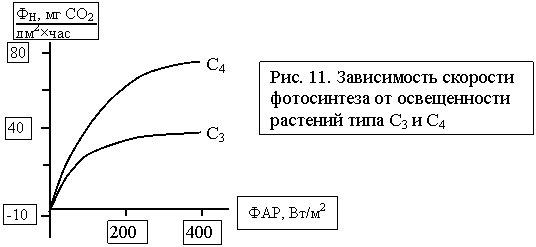
Солнечные лучи
– источник энергии при фотосинтезе, и изменения в скорости фотосинтеза (ФН),
вызываемые меняющейся интенсивностью облучения, относятся к основным
характеристикам фотосинтетической деятельности растений. Зависимость ФН
от облучения выражается характерной кривой (рис. 11), которую обычно называют
световой кривой или кривой освещения: расход СО2
в темноте (при дыхании) с постоянным возрастанием интенсивности освещения
снижается и достигает нулевого значения в так называемой точке компенсации. Дальнейший
рост интенсивности освещения вызывает почти линейное увеличение ФН.
При низких величинах освещения использование лучевой энергии бывает максимальным,
и в таких условиях она служит фактором, лимитирующим ФН. Во второй
части кривой освещения происходит почти полное световое насыщение. В этих
условиях скорость фотосинтеза ограничивается скоростью транспорта СО2 из окружающего растение воздуха в хлоропласты и
скоростью ферментных реакций фиксации СО2. На рисунке 11 приведены
величины ФН, характерные для растений типа С3 и С4.
Углекислота
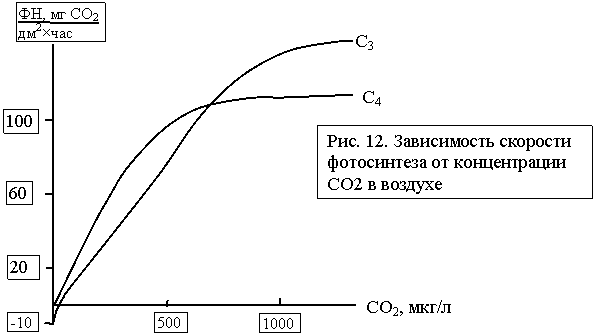
служит субстратом для образования новых продуктов ассимиляции. Концентрация СО2 в воздухе очень мала (0,03%), и каждое ее изменение
отражается на величине ФН.
На рисунке 12,
где приведена зависимость ФН от концентрации СО2,
заметна разница в величине точки компенсации СО2. Кривые освещения
для растений типа С3 и С4 показывают,
что у растений С4 насыщение углекислотой происходит при более низких
ее концентрациях, чем у растений С3. Этот фактор используют на
практике: обогащение атмосферы в оранжереях углекислотой дает существенный
эффект именно у растений типа С3.
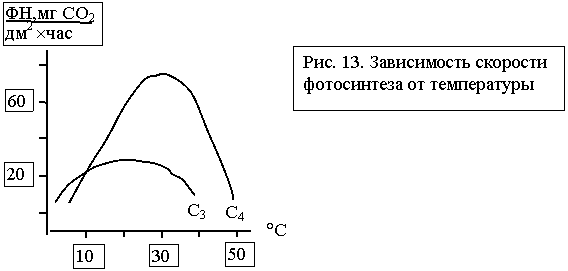
Температура
Температура
оказывает влияние на ход всех ферментных реакций, поэтому отчетливо сказывается
на скорости фотосинтеза (рис. 13). У некоторых видов хвойных деревьев процесс
фотосинтеза протекает и при отрицательных температурах, но у всех полевых
культур прекращается при нескольких градусах выше нуля. На рисунке 13 также
заметно существенное преимущество растений С4
с точки зрения активности фотосинтеза при повышении температур. И наоборот, у растений С3 при одинаковых прочих
условиях скорость фотосинтеза возрастает при более низких температурах. Следовательно,
можно сделать практический вывод: для повышения скорости фотосинтеза растениям
С4 необходимы благоприятные температурные
условия.
Однако работа с кукурузой показывает, что варьирующую зависимость скорости фотосинтеза от температуры можно использовать путем целенаправленной селекции при получении высокопродуктивных сортов для относительно холодных условий. Следует отметить, что успешное возделывание кукурузы в холодных областях не обусловлено лишь взаимосвязью между скоростью фотосинтеза и температурой, хотя именно эту зависимость используют в селекции данной культуры.
Влияние воды
Влияние воды
на скорость фотосинтеза выражено величиной дефицита насыщения, отражающей то
количество воды, которого не хватает растению для полного насыщения.
Максимальной скорости фотосинтеза растения достигают при очень низких
показателях дефицита: при 12-20% устьица заполнены водой с
скорость фотосинтеза падает до нуля (подобные сведения в литературе
[166,190,195]).
Основной
предпосылкой для фотосинтеза является наличие хлорофилла. Его количество
колеблется в пределах 2-6 мг на
Минеральные вещества
Научные
исследования и практика показывают, что на фотосинтез оказывают существенное
влияние минеральные вещества. Дефицит любого из следующих элементов – N, P, K, Mg, S, Ca, Fe, Mn, Cu, B, Zn, Mo, - отрицательно сказывается
на скорости фотосинтеза. Воздействие питательных веществ на фотосинтетический
аппарат бывает многосторонним. Они влияют на формирование морфологической и
анатомической структуры растения, на физические процессы, участвуют в создании
хлоропластов, причем входят в состав как отдельных структурных образования, так
и ферментов.
Определение
количественных характеристик оптимальных условий минерального питания –
довольно трудная задача. Определенное исключение составляет лишь азот. У многих
видов растений установлена зависимость между содержанием азота в ассимилирующих
тканях и скоростью фотосинтеза. Например, показанная на рисунке 14 зависимость
ФН от количества азота в пластинках верхних листьев озимой пшеницы в полевых
условиях. На рисунке также показаны величины обычного содержания азота в
листьях.
ФН 45 3 2

Возраст
Возраст
отдельных органов и растения в целом является существенным эндогенным фактором,
определяющим скорость фотосинтеза. У молодого листа ФН увеличивается до
определенного максимума, достигая его еще перед окончанием роста листа, т.е. до
формирования его полной площади. Этот период называется фотосинтетической
зрелостью листа и длится разное время.
Потенциальная продуктивность фотосинтеза
В связи с значением энергетических сторон фотосинтеза необходимо
обратить внимание и на некоторые другие аспекты данного вопроса. Сравнение
эффективности использования солнечной радиации растениями в широтах России со средними показателями в
мире показывает, в растениеводстве России имеются большие резервы. Для полноты
картины необходимо сопоставление с показателями потенциальной продуктивности
культур, т.е. максимально возможной продукции вещества в оптимальных условиях
внешней среды.
Дальнейшие
примеры иллюстрируют не только возможность получения максимальной продукции, но
и способы ее достижения. Такие методы позволяют анализировать и выявлять
возможные причины значительной разницы между потенциальными (максимально
возможными) и реальными (фактически получаемыми) урожаями в хозяйствах.
Первый пример основан
на механизме фотосинтеза, из которого вытекает, что максимально возможная
эффективность использования ФАР составляет примерно 22%. В условиях Черноземья
сумма радиации за вегетационный период составляет примерно 105´103Дж/см2, что соответствует примерно 47´103 Дж/см2
ФАР (~45% от суммарной солнечной радиации). Это
позволяет синтезировать до 45 т/га сухой массы. Фактические рекордные урожаи в
производственных посевах Кубани достигают 10 т/га сухой массы, т.е. фактическая
продуктивность составляет около четверти потенциальной.
´100 Фиксированная радиация
суммарная радиация
Во
втором примере в опытных условиях получен урожай 15 т/га сухого вещества за 4
месяца. Сумма солнечной радиации за время опыта составила 2,1 млрд. Дж на
![]()
Видно, что данный
высокопродуктивный посев в опыте использовал 1,2% солнечной радиации или примерно
2,5% от ФАР.
Следует
отметить, что в начале ХХ века урожаи зерновых культур колебались в пределах 10
ц/га. Если предположить, что при этом получали
примерно 15 ц/га соломы, то затраты энергии
соответствовали 42,6 млрд. Дж, или 0,2% солнечной радиации. Следовательно,
повышение эффективности использования растениями солнечной радиации с 0,2 до
1,2% сопровождалось ростом общей продуктивности с 25 до 150 ц/га.
Все
приведенные в главе о фотосинтезе факты действуют не разрозненно, а находятся в
довольно сложной взаимосвязи. Например, временный дефицит воды или питательных
веществ может ускорить старение листьев и преждевременное разложение хлорофилла
с последующим падением скорости фотосинтеза. Оценивать это комплексное
взаимодействие можно, лишь разложив его на составные части. Поэтому изучение
основных факторов, влияющих на фотосинтез, очень актуально и необходимо для
оценки состояния посевов с точки зрения формирования ими урожаев в условиях
интенсификации возделывания сельскохозяйственных культур.